
同樣兩個神經元,為什麼「先後順序」差幾毫秒,命運就完全相反?
從 STDP 的毫秒時序、鈣假說與 CaMKII 分子開關,到突觸標記、系統固化與睡眠重播,看可塑性的引擎室如何把短暫訊號刻成永久記憶。
同樣兩個神經元,為什麼「先後順序」差幾毫秒,命運就完全相反?
讀過入門篇的你,已經把「一起激發的神經元會連在一起(cells that fire together, wire together)」這句赫布定律(Hebb's rule)記得滾瓜爛熟。它優雅、好記,也確實抓住了學習的精神。但如果我們把它當成完整的機制來看,會立刻撞上一個尷尬的問題:「一起(together)」到底是多一起?
想像突觸前神經元 A 和突觸後神經元 B。如果 A 在 B 激發「之前」幾毫秒放電,跟 A 在 B 激發「之後」幾毫秒放電,這兩種情境都算「一起激發」嗎?赫布的原始敘述對此語焉不詳。但實驗給出的答案,遠比那句口訣精緻、也殘酷得多:差幾毫秒的先後順序,會決定這個突觸是被增強,還是被削弱——而且兩者的分界,銳利得像一把刀。
這篇進階文章,我們不再重述「大腦會改變」這件事,而是要走進可塑性的引擎室,看清楚三件入門篇刻意略過的事:突觸如何用「時間差」做運算(STDP)、單一個訊號分子如何同時扮演學習的「開關」與記憶的「保險絲」(CaMKII 與鈣假說),以及一條記憶是如何在數小時、數天、數週的尺度上,從脆弱的痕跡被「重新謄寫」成永久檔案(突觸標記、系統固化與睡眠重播)。
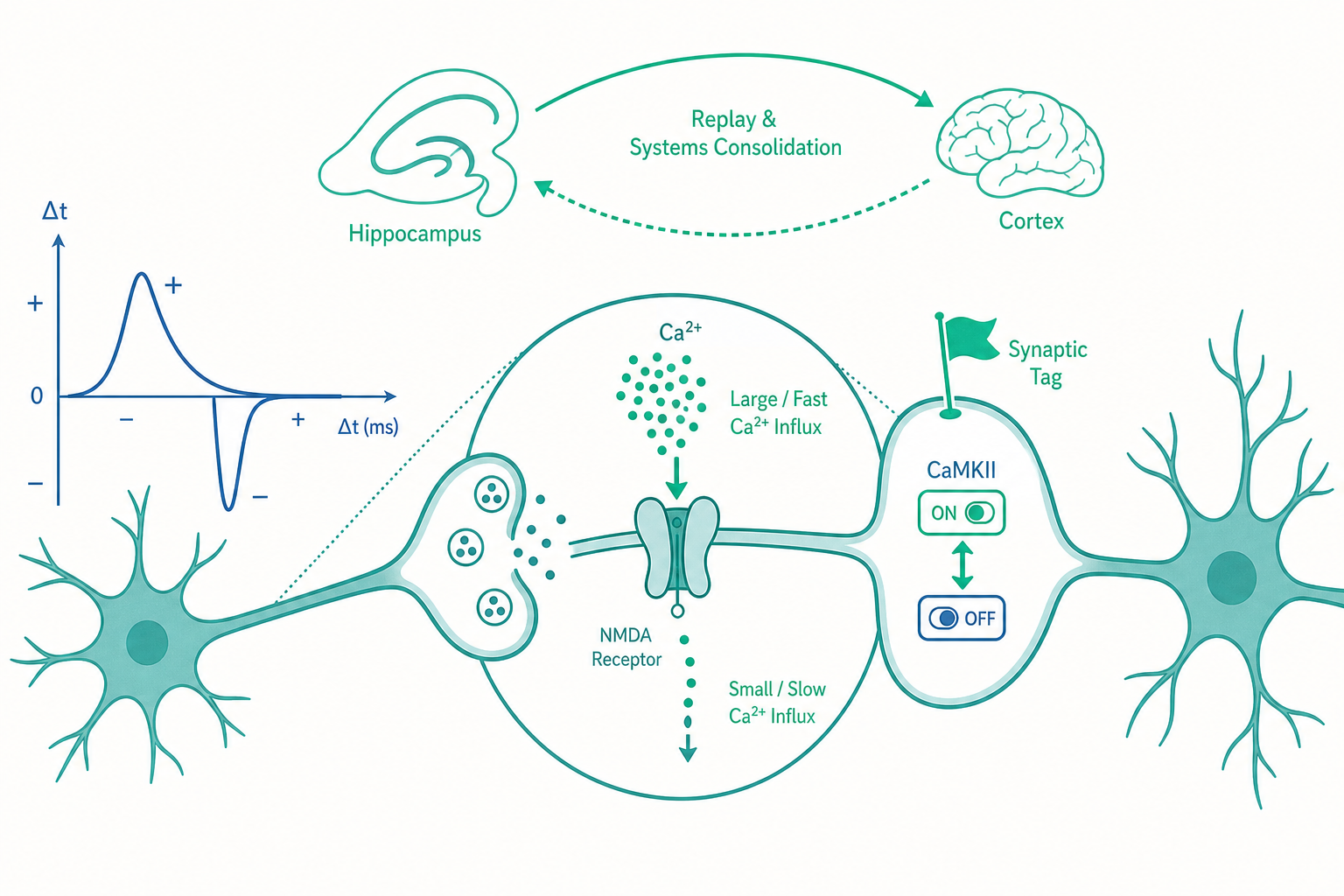
STDP:把赫布定律改寫成一條毫秒級的時序法則
1997 年前後,Henry Markram、Bert Sakmann,以及隨後 Guo-qiang Bi 與 Mu-ming Poo 的一系列實驗,把赫布定律的「一起」精確化到了毫秒尺度。他們的做法是同時記錄突觸前與突觸後兩個神經元,並人為控制它們放電的「相對時間差(Δt)」,然後測量突觸強度怎麼變。結果催生了一個至今仍是可塑性核心的概念——棘波時序依賴可塑性(spike-timing-dependent plasticity, STDP)。
規則出奇地簡潔,可以濃縮成一句因果律:
- 若突觸前神經元「先」放電、突觸後「後」放電(Δt 為正,約 0–20 毫秒內):突觸被增強(LTP)。因為前細胞的放電「有可能參與造成」後細胞放電,符合因果方向,連結被獎勵。
- 若突觸後「先」放電、突觸前「後」放電(Δt 為負):突觸被削弱(LTD)。因為前細胞放得太遲,沒能參與後細胞的激發,這條連結被判定為「不負責任」,遭到懲罰。
把不同 Δt 對應的突觸變化畫成圖,會得到一條著名的「STDP 曲線」:橫軸是時間差、縱軸是突觸強度變化,在 Δt=0 附近呈現一個劇烈的不連續——往正的方向一點點就是強化的尖峰,往負的方向一點點就是削弱的深谷。這正是赫布定律被忽略的下半句(沒一起激發就削弱)在時間軸上的精確展開。
STDP 之所以深刻,在於它讓單一突觸自己就成為一個因果偵測器:它不只問「A 和 B 有沒有同時活動」,而是問「A 的活動是不是 B 活動的『原因』」。這對大腦學習因果關係、預測序列(例如語言中的詞序、音樂中的旋律、運動中的動作鏈)至關重要。一個只懂「同時」的系統無法區分因與果,但一個懂「先後」的系統可以。
看一個例子:海馬迴如何用 STDP 學會「位置序列」
把 STDP 放進一個真實情境會更清楚。老鼠在迷宮裡跑動時,海馬迴中有一群「位置細胞(place cell)」,每個細胞只在動物經過某個特定地點時放電。當老鼠沿著一條路徑跑過 A 點、B 點、C 點,對應的位置細胞會依序激發:A 細胞先、B 細胞次之、C 細胞最後。
按照 STDP 的正向規則,「先放電的 A 細胞」到「後放電的 B 細胞」之間的突觸會被增強;同理 B→C 也被增強。但反方向(C→B、B→A)因為時序顛倒,則被削弱。幾趟下來,整條路徑的「前向連結」被強化成一條單向的神經鏈。
這帶來一個驚人的後果:之後當老鼠只是「站在 A 點」,A 細胞激發就能透過這條被雕刻好的前向鏈,搶先預測性地激活 B、C 細胞。動物因此能在身體還沒走到之前,就在神經層級「預演」接下來的路徑。這正是序列學習與預測的細胞基礎——而它直接源自 STDP 的時序不對稱性。順帶一提,這也是睡眠中「記憶重播(replay)」現象的舞台,我們稍後會回來。
鈣假說:為什麼同一個訊號,既能教 LTP 也能教 LTD?
入門篇告訴你 NMDA 受體是「巧合偵測器」,鈣離子(Ca²⁺)湧入是關鍵。但這裡藏著一個漂亮的謎題:LTP(增強)和 LTD(削弱)這兩個方向相反的結果,用的居然是同一個入口、同一種離子。大腦怎麼可能用同一個訊號分子,既下達「變強」又下達「變弱」兩種相反的指令?
答案是當代可塑性理論最優雅的洞見之一——鈣控制假說(calcium-control hypothesis)。關鍵不在「有沒有鈣」,而在鈣訊號的「振幅」與「時間動力學」:
- 大量、快速、短促的鈣湧入 → 觸發 LTP。這對應 STDP 的正向時序:突觸後剛好強烈去極化時,麩胺酸(glutamate)抵達,NMDA 通道大開,鈣猛灌。
- 少量、緩慢、持續的鈣流入 → 觸發 LTD。這對應弱的、不同步的、或時序顛倒的活動:NMDA 通道只開一條縫,鈣涓涓細流。
換句話說,突觸後細胞讀的不是鈣的「有無」,而是鈣的「劑量與節奏」。高鈣優先啟動激酶(kinase,如 CaMKII),把受體「加上去」;低鈣優先啟動磷酸酶(phosphatase,如 calcineurin / PP1),把受體「拿下來」。同一支溫度計,刻度不同,命運就相反。這也讓 STDP 曲線那條銳利的分界線有了分子解釋:時間差改變了突觸後去極化與麩胺酸抵達的重疊程度,從而改變了鈣的劑量,最終決定走向 LTP 還是 LTD。
CaMKII:學習的開關,也是記憶的「保險絲」
如果要選一個「分子明星」來扮演記憶從訊號變成痕跡的關鍵角色,多數神經科學家會投給 CaMKII(鈣/鈣調素依賴蛋白激酶 II)。它解決了一個讓人頭痛的悖論:鈣訊號只持續幾秒,記憶卻能維持數十年。一個轉瞬即逝的訊號,如何留下持久的印記?
CaMKII 的巧妙之處在於它的自我磷酸化(autophosphorylation)特性。當鈣大量湧入並激活 CaMKII 後,這個酶會「自己把自己磷酸化」,從此即使鈣濃度降回基線,它仍維持在活化狀態——它變成了一個雙穩態的分子開關(bistable molecular switch),可以停留在「開」或「關」兩個狀態,而且能自己維持。
這正是把「短暫訊號」轉成「持久狀態」的機制核心。活化的 CaMKII 會驅動更多 AMPA 受體插入突觸後膜、強化突觸骨架,讓這次增強被「鎖住」。理論上,這種自我維持的開關甚至可以抵抗蛋白質本身的汰換(單個分子被替換,但整個複合體的活化狀態被保留),這也是它被視為記憶「儲存單元」候選的原因之一。
理解 CaMKII,能幫你看穿一個常見迷思:學習不是「資訊被寫進某個靜態硬碟」,而是「一群會自我維持的分子機器被切換到新狀態」。記憶是一種被主動維持的動態平衡,而不是被動的儲存。這個觀念,稍後會在「記憶為什麼會在每次回想時變得脆弱」這件事上派上用場。
後設可塑性:可塑性自己也會變——BCM 與滑動閾值
到這裡你可能會擔心一件事,這也是入門篇在「恆定可塑性」一段點到的隱憂:如果突觸只要符合正向時序就一律增強,那麼最常用的連結會越來越強、搶走所有資源,最後整個網路飽和、失控。大腦如何防止這場正回饋災難?
答案之一是後設可塑性(metaplasticity)——字面意思是「可塑性的可塑性」,也就是「決定突觸該增強還是削弱的那個門檻,本身也會隨經驗移動」。最經典的理論框架是 1982 年提出的 BCM 理論(Bienenstock–Cooper–Munro theory),其核心是一個會滑動的修正閾值(sliding modification threshold, θ_M):
- 當一個神經元「最近整體活動偏高」時,θ_M 會向上滑——意味著要再觸發 LTP 變得更難、觸發 LTD 變得更容易。系統自動踩煞車,避免失控增強。
- 當神經元「最近整體活動偏低」時,θ_M 向下滑——讓突觸更容易被增強。系統自動踩油門,避免迴路沉寂。
這套機制讓大腦在「彈性學習」與「穩定不崩潰」之間自我調節,與入門篇提到的突觸縮放(synaptic scaling)互補:縮放是「整體按比例升降所有突觸」,BCM 滑動閾值則是「動態調整學習新東西的難易門檻」。兩者一個管音量、一個管靈敏度,共同守住穩定與彈性的鋼索。BCM 還有個漂亮的副產品:它能在數學上推導出神經元發展出「選擇性(selectivity)」的傾向,呼應了 Hubel 與 Wiesel 在視覺皮質觀察到的方向選擇性細胞。
突觸標記與捕捉:脆弱的記憶如何「轉正」成永久檔案
我們前面講的 LTP,其實有兩種:撐幾小時就消退的「早期 LTP(early-LTP)」,以及能維持數天、需要合成新蛋白質的「晚期 LTP(late-LTP)」。差別在於:要把一次增強變成持久的結構改變,細胞核必須轉錄基因、製造一批「可塑性相關蛋白(plasticity-related proteins, PRPs)」運送到突觸去「鞏固」它。
但這帶來一個運送難題:細胞核製造的蛋白質會擴散到「整個細胞」,那它怎麼知道該強化哪一個特定突觸?一個神經元有上萬個突觸,剛剛被活化的可能只是其中幾個。
Frey 與 Morris 在 1997 年提出的「突觸標記與捕捉(synaptic tagging and capture, STC)」假說漂亮地解開了這個結。機制是:當某個突觸被強烈活化,它會在「現場」設下一個暫時性的分子標記(tag)——像在門口插一面旗子。隨後細胞核製造、瀰散全細胞的可塑性相關蛋白,會被「插了旗子的突觸」優先捕捉並使用,沒插旗的突觸則拿不到。於是只有被標記的突觸獲得鞏固,轉成晚期 LTP。
這個假說有一個令人拍案的預測(並已被實驗證實):如果一個突觸只被「弱刺激」(只設了標記、卻不足以自己叫細胞核做蛋白),但在時間相近的窗口內,另一個強刺激恰好讓細胞核大量製造了蛋白質,那麼這個弱突觸竟然能「搭便車」捕捉到那批蛋白,把原本撐不久的弱記憶升級成持久記憶。
這正是行為層面「行為標記(behavioral tagging)」的細胞基礎,也解釋了一個你或許有過的經驗:一件本來微不足道、轉眼會忘的小事,若剛好發生在一個情緒強烈或高度新奇的時刻附近,反而被牢牢記住了。那個強烈時刻啟動了蛋白質工廠,而你那件小事的突觸標記,搭上了這班便車。
系統固化與睡眠重播:記憶會「換地方住」
最後一塊拼圖,是把尺度從突觸(毫秒到小時)拉到整個腦系統(天到年)。入門篇提到記憶固化(consolidation)與睡眠的重要,這裡我們看它的機制版本——系統固化(systems consolidation)。
新的事件記憶(episodic memory)一開始高度依賴海馬迴(hippocampus)。但隨著時間,這份記憶會逐漸轉移、分散到大腦皮質(neocortex)成為相對獨立、不再需要海馬迴的長期記憶。這個「換地方住」的過程,很大程度發生在睡眠中。
關鍵機制是前面埋下的伏筆——記憶重播(memory replay)。在慢波睡眠(slow-wave sleep)期間,白天清醒時被 STDP 雕刻出來的位置細胞序列(還記得 A→B→C 的前向鏈嗎?)會被自發地、加速地重新激發一遍,而且常是壓縮過、甚至倒序的。這場「夜間重播」不只是回放,它伴隨海馬迴的「尖波漣漪(sharp-wave ripple)」與皮質慢波的精細時間耦合,被認為正是把記憶從海馬迴「謄寫」到皮質的搬運工。實驗上若選擇性破壞睡眠中的尖波漣漪,動物的記憶固化就會受損。
這也解釋了入門篇「睡眠對學習關鍵」這句話背後的硬機制:你睡覺時,大腦並沒有閒著,它在用 STDP 與重播,把白天脆弱、依賴海馬迴的痕跡,一筆筆轉謄成穩固、分散在皮質的長期檔案。睡眠不是學習的休止符,而是固化的主場。
重點回顧
- STDP 把赫布定律精確化到毫秒:突觸前先放電→增強(LTP),突觸後先放電→削弱(LTD),分界銳利。它讓單一突觸成為偵測「因果方向」的元件,是序列與預測學習的基礎。
- 鈣假說解釋了一個入口兩種結果:不是鈣的有無,而是鈣的「振幅與節奏」——大量快速的鈣走 LTP(激酶主導),少量持續的鈣走 LTD(磷酸酶主導)。
- CaMKII 是雙穩態分子開關:透過自我磷酸化把「幾秒的鈣訊號」轉成「可自我維持的活化狀態」,是把短暫訊號變成持久記憶的核心機制。
- 後設可塑性守住穩定與彈性:BCM 的滑動閾值讓「學習難易門檻」隨整體活動動態調整,與突觸縮放互補,防止正回饋失控。
- STC 與系統固化是記憶「轉正」的流程:突觸標記讓特定突觸捕捉細胞核製造的蛋白而被鞏固;睡眠中的重播則把記憶從海馬迴搬遷到皮質,完成長期化。
深入探討(研究所視角)
記憶提取的悖論:再固化(reconsolidation)與「記憶會被改寫」
CaMKII 那節埋下的伏筆在此引爆。一個長期記憶並非取出來看完就原封不動放回——當你「提取(retrieve)」一段已固化的記憶,它會短暫回到「不穩定、可被修改」的脆弱狀態,必須再經一次蛋白質合成依賴的過程才能重新穩固,這就是再固化。這意味著每一次回想,都可能(在分子層級)悄悄改寫那段記憶。這既是創傷後壓力症候群(PTSD)治療的潛在切入點(在提取窗口內介入,弱化恐懼記憶的再固化),也是法律上「目擊證詞為何不可靠」的神經科學解釋。它徹底顛覆了「記憶是錄影帶」的直覺:記憶更像一份每次打開就會被重新存檔、且存檔時可能夾帶當下狀態的活文件。這條線索直接連向優心理的記憶與證詞研究。
工程化可塑性:光遺傳學與「記憶痕跡(engram)」的因果操弄
過去可塑性研究多是「相關性」證據——看到學習伴隨 LTP。近十年,Susumu Tonegawa 等人結合光遺傳學(optogenetics)與活動依賴標記技術,首度做到「因果」操弄:標記出一段特定記憶所對應的神經元集群(engram cells),然後用光「人為地」重新激活它們,在動物身上憑空喚起那段記憶;甚至能植入一段「從未真實發生過」的假記憶。這把可塑性研究從觀察推進到工程,也讓「記憶痕跡」從理論假設變成可被定位、可被操弄的物理實體。其倫理意涵——記憶可被人為植入或抹除——直接呼應 Educational Omics 框架中的 Ethicomics 維度。
跨領域連結:從 EWC 看大腦如何解決 AI 的「災難性遺忘」
入門篇提到人工神經網路(artificial neural network)的災難性遺忘(catastrophic forgetting):學新任務時抹掉舊任務。值得注意的是,機器學習近年最受矚目的解方之一——彈性權重固化(Elastic Weight Consolidation, EWC)——其靈感正是直接來自本文的生物機制。EWC 的做法是:估計每個權重對「舊任務」有多重要,然後在學新任務時,對「重要權重」施加更強的修改阻力,讓它們變「硬」、不易被覆寫——這幾乎就是突觸標記與系統固化的工程版翻譯:把該保留的連結「鞏固住」,把不重要的留給新學習。生物大腦用 STC、滑動閾值、睡眠重播等多重機制天然地實現了「該記的鎖住、該學的開放」;AI 工程師則在反向重新發明這些原則。這正是 Uedu 在 AI 與 優神經科學 之間架橋的核心旨趣:生物可塑性既是 AI 持續學習(continual learning)的靈感泉源,也是一面照出人工系統缺陷的鏡子。
對學習實踐的進階啟示
把這些深層機制收束回學習現場,會得到比入門篇更精準的處方。其一,提取練習(retrieval practice)勝過反覆閱讀——因為提取會觸發再固化,每次成功回想都在強化並更新那條痕跡,而被動重讀只是讓資訊「看起來熟悉」卻沒走過鞏固流程。其二,間隔效應(spacing effect)有了機制解釋——分散練習讓每次學習都落在不同的後設可塑性狀態(滑動閾值在不同位置),並給系統固化留出時間,遠勝塞在一起的集中練習。其三,情緒與新奇是可塑性的放大器——行為標記告訴我們,把要記的內容跟一個有意義、情緒投入或新奇的情境綁在一起,能讓它「搭上」蛋白質合成的便車而更持久。你的大腦終身可被雕塑,但這把刻刀的鋒利程度,取決於你用對了時序、提取、間隔與投入——而這些,全都寫在突觸的分子細節裡。