
如果皮質各區看起來幾乎一模一樣,為什麼有的負責看、有的負責想?
從六層皮質、典型微迴路到預測編碼與連結組,看大腦如何用同一套「運算藍圖」支撐截然不同的認知功能。
如果皮質各區看起來幾乎一模一樣,為什麼有的負責看、有的負責想?
你在入門篇學會了大腦的「地理」:四大腦葉各司其職,腦區分工但不孤立。但這裡藏著一個讓神經科學家困惑了一個世紀的悖論——如果你在顯微鏡下切一小片初級視覺皮質(V1)和一小片前額葉皮質(PFC),它們的細胞構造看起來幾乎一樣:同樣的六層結構、同樣的神經元類型、同樣的連線邏輯。可是一個負責「看見邊緣」,另一個負責「規劃下週的考試進度」。
這不是巧合,而是一個深刻的線索。1978 年,神經科學家 Vernon Mountcastle 提出了一個大膽的假說:皮質之所以能用「同一套硬體」處理視覺、聽覺、語言、決策等天差地別的任務,是因為它演化出一個可重複、模組化的運算單元——他稱之為皮質柱(cortical column)。理解這個「皮質的基本運算模組」,以及它如何透過分層(lamination)與連結組(connectome)串成階層式網絡,是從「背腦區名稱」躍升到「理解大腦如何運算」的關鍵一步。
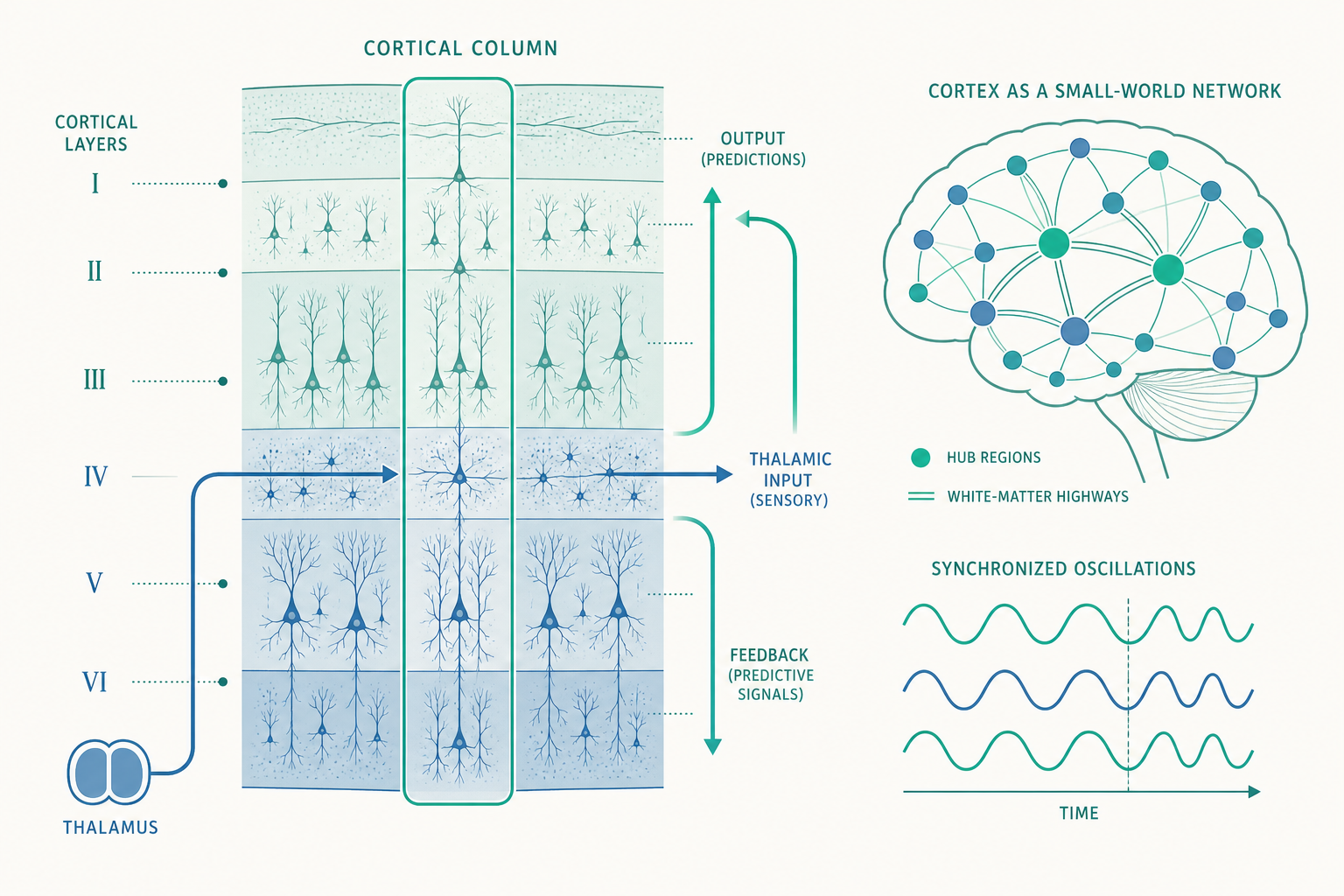
六層皮質:一張被反覆使用的運算藍圖
入門篇把大腦皮質(cerebral cortex)描述成「最外層那片佈滿皺褶的灰質」。但若你垂直切過皮質、從表面往深處看,會發現它並非均質的一團,而是分成六個界線分明的細胞層(layers I–VI),這就是所謂的新皮質(neocortex)六層結構。每一層的細胞種類、密度與連線目標都不同,構成一條精密的訊息「流水線」:
- 第 IV 層(layer IV) 是主要的輸入層:來自視丘(thalamus)的感覺訊號優先抵達這裡。在感覺區(如 V1),第 IV 層特別厚;而在運動皮質這種「以輸出為主」的區域,第 IV 層幾乎消失——這種差異稱為細胞構築(cytoarchitecture) 的區域變異,正是 Brodmann 在 1909 年據以把皮質劃分成 52 個分區的依據。
- 第 II/III 層(上層,supragranular) 主要負責皮質與皮質之間的水平連結,把處理過的訊號送往同階或更高階的腦區。
- 第 V 層與第 VI 層(下層,infragranular) 是主要的輸出層:第 V 層的大型錐體細胞(pyramidal neuron)把指令送往皮質下構造(如基底核、腦幹、脊髓),第 VI 層則送回視丘形成回饋迴路。
請注意這個關鍵洞見:訊息在皮質裡不是隨機亂竄,而是沿著「層」定向流動——輸入進第 IV 層、在柱內垂直整合、再經上層往外送或經下層往下送。Mountcastle 與後來的 Rodney Douglas、Kevan Martin 把這條反覆出現的連線模式抽象成典型微迴路(canonical microcircuit):一個彷彿被大自然「複製貼上」到整片皮質的標準電路圖。視覺區用它分析光點,聽覺區用它分析聲波,前額葉用它操作抽象概念——硬體相同,差別只在於輸入的內容和與誰相連。
皮質柱:垂直方向的功能模組
如果說「層」是水平的分工,那麼皮質柱就是垂直方向的功能單元。Mountcastle 在研究體感覺皮質時發現:當電極垂直插入皮質、由表面往深處走,沿途遇到的神經元對同一種刺激有反應(例如同一根手指的觸覺);但只要電極稍微傾斜、跨越到隔壁的柱,反應的偏好就突然改變。
最經典的證據來自 Hubel 與 Wiesel 對視覺皮質的研究(他們因此獲得 1981 年諾貝爾生理醫學獎)。他們發現 V1 中的神經元各自偏好特定方向的線條——有的對垂直線反應最強,有的偏好 45 度斜線。而這些「方向偏好」在皮質表面有系統地平滑變化,形成所謂的「方向柱(orientation column)」與「眼優勢柱(ocular dominance column)」交織的優美地圖。
這帶來一個重要的觀念修正:入門篇說「腦區分工」,進階版要補上——分工是有層級尺度的。從巨觀的腦葉,到中觀的功能分區(如 V1、V2、V4),再到微觀的皮質柱(直徑約 0.3–0.5 毫米),大腦在每一個尺度上都展現模組化。這種「巢狀模組(nested modularity)」正是它既能高度專精、又能彈性整合的結構祕密。
看一個例子:一條斜線如何在皮質裡被「算」出來
讓我們把上面的概念落地,看 V1 如何偵測一條傾斜的邊緣:
- 視丘輸入分散又單調:來自外側膝狀體(LGN,視丘的一部分)的神經元只會報告「這個小點亮、那個小點暗」,它們的感受野(receptive field)是一個個圓形的亮暗對比,本身完全不知道方向。
- 第 IV 層做空間整合:Hubel 與 Wiesel 提出的經典模型認為,若把一排沿斜線排列的 LGN 輸入,全部匯聚到同一個 V1 神經元的第 IV 層樹突上——那麼這個 V1 神經元就只在「該斜線上同時有光」時才強烈激發。換句話說,方向選擇性不是天上掉下來的,而是由下層輸入的幾何排列『算』出來的。
- 柱內垂直放大與銳化:訊號在皮質柱內垂直傳遞,經由抑制性中間神經元(interneuron)的側向抑制(lateral inhibition),把「最符合的方向」放大、把「接近但不完美的方向」壓下去,使方向調諧曲線變得更尖銳。
- 上層往外送、形成階層:處理過的「這裡有一條 45 度邊緣」訊息,經第 II/III 層送往 V2、V4,在那裡與其他邊緣組合成角、輪廓,最終在顳葉的高階視覺區拼成「這是一張臉」。
整條流水線的精妙之處在於:每一階都用同一套微迴路,只是把上一階的輸出當成自己的輸入。複雜的知覺,是簡單運算單元一層層堆疊的湧現結果。
階層、回饋與預測編碼:大腦不只是被動接收
入門篇強調訊號「由下而上(bottom-up)」從感覺器官送往皮質。但進階的真相是:回饋連結(feedback)的數量往往比前饋(feedforward)還多。從高階區送回低階區的軸突,數量驚人。大腦為什麼要花這麼多資源「往回傳」?
當代一個極具影響力的框架是預測編碼(predictive coding),由 Rao 與 Ballard(1999)等人系統化提出。核心想法是:大腦不是被動等待感覺訊號,而是不斷主動產生對世界的預測,然後只把「預測錯誤(prediction error)」往上傳。
- 高階腦區透過回饋連結,向低階區「下注」:預測接下來會看到/聽到什麼。
- 低階區把實際輸入與這個預測相比,只把不符之處(誤差)往上報告。
- 上層據此更新模型,再下達新的預測,如此往復收斂。
這個框架優雅地解釋了許多現象:為什麼你能「看見」眼角餘光中其實模糊的東西(大腦用預測補完)、為什麼錯字常被自動忽略(符合預測就不報錯)、為什麼意料之外的事件會引發強烈的腦反應(巨大的預測誤差)。它也把感知、注意力、學習統一在一個數學框架下——學習,就是最小化長期的預測誤差。
值得注意的是,這個觀念與當代 AI 深有呼應:在優 AI 的視角下,自監督學習(self-supervised learning)中的「預測下一個詞元(token)」或「還原被遮蔽的影像區塊」,本質上正是讓模型透過最小化預測誤差來建構世界模型——這與預測編碼的精神不謀而合。但同樣要謹慎:生物大腦的預測編碼是否真的以反向傳播(backpropagation)那樣的方式更新突觸,至今仍是開放的爭論。
白質連結組:腦區之間的「高速公路」
入門篇談的是灰質(gray matter,神經元細胞本體聚集處)。但皮質之所以能組成網絡,靠的是底下的白質(white matter)——由覆著髓鞘(myelin)、外觀偏白的軸突束組成的「纜線」。理解大腦的網絡,就必須理解這些連線。
借助擴散張量造影(diffusion tensor imaging, DTI) 與纖維追蹤(tractography)技術,研究者得以非侵入地描繪出活人大腦的主要纖維束,例如連接額葉與顳葉、對語言至關重要的弓狀束(arcuate fasciculus)。把所有腦區當成節點、纖維束當成連線,繪製出的整體連線圖就是連結組(connectome)。
連結組研究揭示了大腦網絡的幾個深刻特性:
- 小世界網絡(small-world network):大腦同時擁有「密集的局部連結」與「少數的長程捷徑」,使得任意兩個腦區之間平均只需經過很少的步數就能溝通——既節省連線成本,又保有高效整合。
- 樞紐(hub)與富人俱樂部(rich club):少數高度連結的核心腦區(如後扣帶迴、楔前葉)彼此之間連結特別緊密,形成一個「富人俱樂部」,承擔了跨網絡整合的重任。也正因如此,這些樞紐一旦受損(如某些中風或退化性疾病),影響往往特別廣泛。
- 整合與分離的權衡(integration vs. segregation):健康的認知,需要在「模組各自獨立運算」與「全腦資訊整合」之間取得動態平衡。許多精神與神經疾病,被重新理解為這種網絡平衡的失調,而非單一腦區的損壞。
神經振盪:用「節奏」綁定分散的訊息
最後一塊進階拼圖回答了一個入門篇沒處理的難題:如果一個物體的「顏色」在某個腦區處理、「形狀」在另一個腦區、「動作」又在第三個腦區——大腦怎麼知道這些分散的特徵屬於『同一個』物體? 這就是著名的「綁定問題(binding problem)」。
一個重要的候選答案是神經振盪(neural oscillation),也就是腦電圖(EEG)上看到的各種頻段腦波。當分散在不同腦區的神經元群以同步的節奏一起放電時,它們彷彿在說「我們是一夥的」。
- 伽馬波(gamma,約 30–100 Hz) 被認為與局部的特徵綁定、注意力聚焦有關。
- theta 波(約 4–8 Hz) 與海馬迴的記憶編碼、空間導航密切相關。
- 不同頻段之間還存在跨頻耦合(cross-frequency coupling),例如慢波的相位調控快波的振幅,被視為大腦協調不同時空尺度運算的機制。
這帶出一個與入門篇互補的觀點:腦區之間不只靠「解剖上連不連」溝通,更靠動態的時間協調。同一套解剖連線,可以透過不同的同步模式,靈活地組成不同的「功能網絡」——這正是大腦在固定硬體上實現彈性運算的另一層祕密。
重點回顧
- 皮質的強大來自可重複的運算模組:六層結構與典型微迴路(canonical microcircuit)讓視覺、聽覺、決策能共用一套「硬體藍圖」,差別只在輸入與連線對象。
- 模組化是多尺度、巢狀的:從腦葉、功能分區到皮質柱(cortical column),大腦在每個尺度都展現分工,方向選擇性等特性是由下層輸入的幾何排列「算」出來的。
- 大腦是主動的預測機器:回饋連結往往多於前饋,預測編碼(predictive coding)主張大腦只把「預測誤差」往上傳,學習即是最小化長期誤差。
- 白質連結組決定網絡拓撲:大腦呈現小世界與富人俱樂部特性,認知健康仰賴整合與分離的動態平衡,樞紐受損影響特別廣泛。
- 神經振盪提供時間維度的協調:透過同步節奏(gamma、theta 等)綁定分散的特徵,同一套解剖連線可組成不同的功能網絡。
深入探討(研究所視角)
走到這裡,幾個前沿議題與爭論值得你放進雷達。
皮質柱:是真實的解剖實體,還是有用的抽象? Mountcastle 的皮質柱假說影響深遠,但近年也面臨嚴肅質疑。批評者(如 Jonathan Horton 與 Daniel Adams)指出,「柱」的界線在不同物種、不同皮質區並不一致,有些區域根本找不到明確的柱狀構造,因此「皮質柱是皮質的普遍基本單元」這個強主張可能過度推廣。較穩健的當代立場是:把柱視為一種有用的功能組織原則與分析尺度,而非處處存在的剛性解剖磚塊。這個爭論本身就是科學的好教材——一個漂亮的理論如何在更多資料下被修正、限定其適用範圍。
從「典型微迴路」到細胞類型的精細圖譜。 近年由 Allen Institute 等主導的大規模計畫,結合單細胞 RNA 定序(single-cell RNA-seq)、形態學與電生理,正在繪製前所未有精細的皮質細胞類型圖譜(cell-type atlas),揭示出遠比「興奮性錐體細胞 vs. 抑制性中間神經元」二分法複雜的多樣性。這提醒我們:典型微迴路是一個第一近似,真實迴路的計算能力,深植於這些細胞類型各自的分子與生理特性之中——這正與優生物的分子神經科學接軌。
全腦模型與連結組學的計算化。 把連結組(結構連線)與神經振盪(功能動態)結合,研究者開始建構全腦計算模型,用一組耦合的微分方程模擬整片皮質的活動,探討為何特定的結構拓撲會湧現出特定的功能網絡(如預設模式網絡)。這是圖論、動態系統理論與神經科學的交會點,也是計算神經科學最活躍的前沿之一。
與人工智慧的雙向對話,需保持清醒的類比。 深度學習中的卷積神經網路(CNN)其階層式特徵抽取,與 Hubel-Wiesel 的視覺階層驚人地相似;Transformer 的注意力機制,也常被拿來與大腦的選擇性注意對照。在優 AI 與優心理的交叉點上,這些類比極富啟發性。但研究所階段更重要的是辨識類比的邊界:真實皮質有大量回饋連結、稀疏放電、神經調質(neuromodulator)動態調控與能量極度節約等特性,是當前主流人工網路所缺乏的。預測編碼、神經形態運算(neuromorphic computing)與類腦的局部學習法則(local learning rules),正是試圖把這些生物洞見帶回 AI 的努力方向。
把這些線索接起來,你會看到「大腦解剖」在進階層次的真正面貌:它不是一張靜態的腦區地圖,而是一個多尺度的、會主動預測的、靠節奏協調的運算系統。從六層微迴路到全腦連結組,從預測誤差到伽馬同步,這些正是神經科學、生物學、心理學與人工智慧在二十一世紀共同追問的核心——結構如何生成計算,計算又如何成就心智。